LÄHDE: Paues Jakob. Brain Stem Involvment in Immune and Aversive Challenge. Linköping universitet. 2006 ISBN 91-85643-81-5
Poimintoja abstraktista ja taustateoriasta suomeksi.
Bakteerien ym vaikuttama immuunijärjestelmän aktivaatio aiheuttaa akuuttifaasivasteen ja sairaudelle tyypillisen käyttäytymisen, kun aivot koordinoivat autonomisia, endokriinisiä ja käytöksellisiä vasteitaan. Jälkimmäiset muodostuvat KUUMEEN, VÄSYMYKSEN, ANOREXIAN ja HYPERALGESIAN vaikutuksista. Usein tieteessä käytetty sairauskäyttäytymisen malli saadaan koe-eläimessä aikaan iv- injektiolla, jossa annetaan bakteeriendotoksiinia, lipopolysakkaridia (LPS). LPS aiheuttaa inflammatoristen välittäjäaineitten tuoton. Näitä ovat SYTOKIINIT ja PROSTAGLANDIINIT, ja ne puolestaan voivat vuorovaikuttaa keskushermostoon (CNS) ja sitä kautta sairaudelle tyypillistä käyttäytymistä. Ihmisellä on keskushermostossa myös muistikuva niistä substansseista, jotka ovat aiemmin tehneet sairaaksi ja muistikuvan tarkoituksena on pohjustaa tulevaisuudessa tapahtuvan vahingon välttämistä. a tämän ilmiön nimenä on ”conditioned taste aversion” (CTA) ehdollinen makuaistimuksen aversio ( torjunta) Usein käytetty koe-eläinmalli tämän ehdollistuneen torjunnan, CTA, tutkimukseen on intraperitoneaalinen injektio LiCl, lithiumkloridia.
AISTIMUKSIEN radat periferiasta aivokeskukseen kulkevat afferenttina (aivoihin päin nousevana) somatosensorisena ja visceroseptiivinä informaationa, mikä tieto integroidaan aivorungon rostraalisessa osassa aivosillan (pons) haarojen ( brachium) parabrachiaalisissa tumakkeissa (PB), joissa sijaitsee autonominen reletumake ja siitä tieto välittyy edelleen ylemmälle tasolle useisiin hypotalamuksen ja etuaivojen (limbisiin) rakenteisiin.
KUVA 1. Pons, aivosilta http://i.ytimg.com/vi/XN6NzyleE2k/0.jpg
KUVA 2. Parabrachiaalinen nukleus PB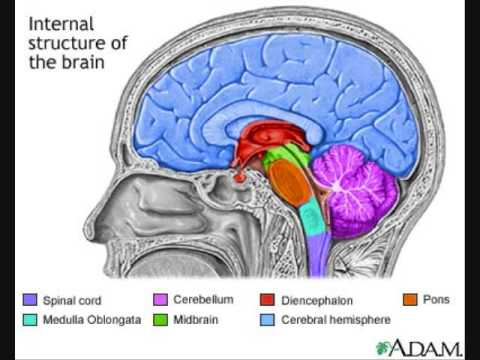{kind=link}
http://www.georgiapainphysicians.com/downloads/m1_slides/12.%20opioid%20receptors.jpg
{kind=link}
AIVOSILLAN (pons) parabrachiaaliset tumakkeet (PB) ovat kriittisen olennaisia, jotta voi kehkeytyä ehdollisen makuaistimuksen aversiota, torjuntaa (CTA) ja parabrachiaaliset neuronit aktivoituvat monella anorexigeenillä aineella.
Lisäksi nämä parabrachiaaliset neuronit (PB) ilmentävät NEUROPEPTIDEJÄ joista mainitaan KALKITONIINI-geeniin liittyvä peptidi CGRP (calcitonin gene related peptide) ja ENKEFALIINI (encephalin). Näiden molempien on havaittu osallistuvan immunoisgnalointiin, nosiseptioon, ravinnon ottoon ja aversioon.
Työn ensimmäinen osa (I) Paues J et al. Preproencephalin mRNA expression in rat parabrachial neurons; relation to cells activated by systemic immune challenge. Neurosci Lett 2001; 316; 165-8.
Tässä väitöskirjatyön osassa tutkittiin enkefalinergisiä neuroneita parabrachiaalisesta tumakkeesta (PB). Niitä aktivoitiin systeemiseen immunologiseen vasteeseen. Tätä vastetta mitattiin immunohistokemiallisesti in situ- hybridisaatiotekniikalla kaksoismerkkausta käyttäen. Moni neuroni kyllä ilmensi heti varhaista fos-geeniä (bakteerisubstanssin) LPS- injektion jälkeen niistä hermosoluista, jotka sijaitsivat parabrakiaalisen tumakkeen (PB) uloimmassa reunassa (PBel; external lateral parabrachial subnucleus). Suuri osa näistä PBel- neuroneista ilmensi PRE-PRO-ENKEFALIINIA. Mutta kaksoismerkattuja soluja oli vain vähän. Fos-geeniä ilmeni niissä soluissa, jotka olivat ulommassa osassa ( outer part) PBel tumaketta ja merkattiin sen takia PBelo- hermosoluiksi. Sen sijaan PRE-PRO-ENKEFALIINIA ilmentävät PBel-neuronit sijaitsivat lähimpänä pedunculusta. Tästä tiedemiehet päättelivät, että vaikka ENKEFALIINI mainitaan autonomisen ja immuunisignaloinnin osatekijänä, PB-ENKEFALIINI-ergiset neuronit eivät näytä kuitenkaan aktivoituvan immuunistimuluksesta.
Työn toinen osa (II) Paues J et al. Feeding related immunoresponsive brain stem neurons: association with CGRP. Neuroreport; 2001; 12; 2399-403
Jotta saataisiin edelleen luonnehdittua PBelo- neuroneitten immuunialtistuksestta johtuvaa aktivoitumista, katsottiin ilmensivätkö nämä neuronit CGRP:tä, (Calcitonin gene related peptide). Kaksoismerkkaus ja in situ hybridisointi osoittivat, että ne PBelo-neuronit, joissa ilmeni fos-geeni heti bakteeriperäisen LPS-materiaalin injektion jälkeen, suurelta osaltaan ilmensivät samanaikaiseti CGRP mRNA:ta, mikä viittaa siihen, että CGRP saattaa olla osallisena immunologiseen altistukseen liittyvässä sairausvasteessa ja aiheuttaa sytokiinien indusoimaa anorexiaa.
Työn kolmas osa (III) Paues J et al. Activation of the parabrachio-amygdaloid pathway by immune challenge or spinal nociceptive input: a quantitative study in the rat using Fos immunohistochemistry and retrograde tract tracing. J Comp Neurol 2005
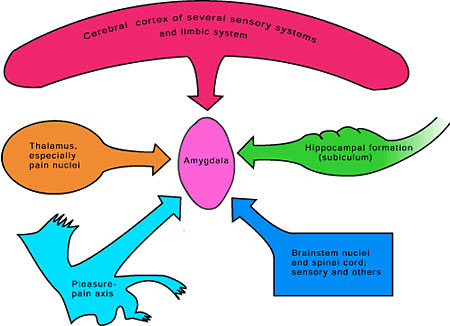
KUVA 3-4.Mantelitumake, amygdala
http://www.drawittoknowit.com/radiographic-atlas/amygdaloid-body.html
http://www.neilslade.com/gifs/AmygdalaInputs.jpg
{kind=link}
Kaksoismerkkauksella ja immunohistokemiallisesti tutkittiin, jos ne PBel-neuronit, jotka aktivoituivat immuunistimuluksesta, omasivat projektiota amygdalaan (aivojen mantelitumakkeeseen), joka on limbinen rakenne ja joka osallistuu homeostaasissa tapahtuvaan altistuksen affektiiviseen vasteeseen.
Koe-eläimiin injisoitiin amygdalaan retrogradinen merkkausaine (tracer) koleratoksiini b (CTb) ja sen jälkeen immuunialtistettiin. Tutkijat totesivat, että LPS-substanssia jälkeen Fos-geeniä heti ilmentäneistä neuroneista kolmasosa merkauttui myös CTb:llä. Tästä nähtiin, että immuunialtistuksesta aktivoituneet PBe- neuronit projisoituivat amygdalaan.
Aimmin on viitattu siihen, että PBel-neuroneilla on nosiseptiivissä ( kipu-) signaloinnissa tärkeää osuutta. Tutkijat halusivat tiettää, jos amygdalaan projisoituvat PBel-neuronit aktivoituvat nosiseptiivistä (kipu-) stimuluksesta ja tässä tarkoituksessa edelleen injisoitiin amygdalaan CTb:tä merkkausta varten. Toipumisen jälkeen koe-eläimen takajalkaan asetettiin formaliinia, mistä tuli kipuimpulssia. Kaksoismerkattu immunohistokemiallinen tutkimus Fos ja CTb:tä kohtaan osoitti, että vain hyvin harva traumalla aktivoitu PB-neuroni näytti projisoituvan amygdalaan. Niinpä PBel-amygdala projektio vaikuttaa olevan tärkeä vain immuunialtistuksessa, mutta ei sen sijaan (nosiseptiivissä) kipusignaloinnissa
(Nosiseptiivinen eli kipusignalointi käytti rataa sisemmän lateraalitumakkeen puolella, jonka projektio on intralaminaariseen talamukseen ja ventrolateraaliseen ydinjatkokseen ja selkäytimeen) Siis iso määrä PBel neuroneista ei vastaa immuunistimuluksiin ja sen sijaan immuunistimuluksista aktivoitii PBel kohdassa vain CGRP-ergiset neuronit.
KUVA 5. Näkökukkula, thalamus , synhögen
http://sv.wikipedia.org/wiki/TalamusTyön neljäs osa (IV) Paues J et al. Expression of melanocortin- 4 receptor by rat parabrachial neurons responsive to immune and aversive stimuli. Neuroscience 2006; 141; 287-97.
Moni PBel-neuroni ilmensi Fos-geenituotetta heti intraperitoneaalisen LiCl-injektion jälkeen.
MELANOKORTIINIT ovat neuropeptideitä, joillä on äskettäin havaittu olevan osuutta aineenvaihdunnassa, ravinnon otossa ja aversiivisissä mekanismeissa. Parabrachiaalisesta tumakkeesta (PB) tiedetään, että siinä sijaitsee MELANOKORTIINI-RESEPTORI-4 (MC-4) mRNA. Yllä mainitulla tekniikalla tutkittiin, jos nämä PB-neuronit jotka aktivoituvat iv injektiosta (LPS) tai intraperitoneaalista injektiosta (LiCl), ilmensivät MC4-R lähetti-RNA:ta. Tutkijat totesivat, että monet tällaiset jommasta kummasta aineesta aktivoituvat PBelo- neuronit ilmensivät suurelta osaltaan MC-4 R mRNA:ta. Jälleen käytettäessä samanlaista merkkaustekniikka ja menetelmää MC-4R mRNA:ta ja CGRPmRNA:tä kohtaan, havaittiin, että iso osa CGRPpositiivisista PBelo-neuroneista ilmensi myös MC-4R mRNA:ta. ( MELANOKORTIINI-reseptorin lähettiRNA:ta).
YHTEENVETONA tästä väitöskirjasta tutkija mainitsee , että perifeerinen immuunialtistus aktivoi PBel-alueen CGRP-neuroneita; lipopolysakkaridilla (LPS) aktivoidut PBel neuronit projisoituvat amygdalaan; amygdalaan projisoituvat neuronit PBel- alatumakkeessa ovat CGRP-positiivisia; immuunialtistuksella tai aversiivilla altistuksella aktivoidut PBel- neuronit ilmentävät MC4-R, MELANOKORTIINI-4 reseptoria.
JOHTOPÄÄTÖKSENÄ nämä tiedot viittaavat siihen, että on olemassa MELANOKORTIINILLA säätyvä CGRP-positiivinen tie aivosillasta, sen aivosillan parabrachiaalisesta lateraalisesta tumakealueesta (PBel) limbisen järjestelmän amygdalaan ( mantelitumakkeeseen) ja tämä reitti (PBelo-amygdala-reitti) toimii releenä sairauteen liittyvän käyttäytymisen tietyissä aspekteissa ( negativ hedonic afferent signalling) antaen perustavaa informaatiota, jolle kehkeytetään affektiivinen vaste sairauteen ja sairauden tunteeseen.
Tätä tietä pitää normaalisti inhibitiossa kehon opioiderginen PB-neuronijärjestelmä. ENKEFALIINIT muodostavat osan anti-nosiseptiivistä (kivunvastaisesta) endogeenisestä systeemistä, jossa on PBeli-PBelo interaktiota tumakkeen alaosien kesken ja siitä seuraa normaalisti ylläpidetty toninen inhibitorinen vaikutus PBelo-neuronien –amygdala projektiin päin.(Tämän basaalitonuksen voi voittaa ”epämiellyttävä” (negativ hedonic) informaatio, mille sitten homeostaasin yläpitämiseksi kehkeytyy vasteen affektiivinen komponentti)
Väitöskirjan taustateoriasta poimintoja homeostaasista, joka on tavallaan biologinen vastine yhteiskunnan poliittiselle demokratialle.
HOMEOSTAASI Keholle on tärkeää että kaikki kehon solut omaavat yhteistoimintaa siten, että kehon sisäisessä miljöössä vallitsee stabiili ja optimaalinen sisäinen miljöö. Tätä prosessia kutsutaan homeostaasiksi ja se on hyvin tehokas keino ihmisen pysymisessä hengissä, vaikka ulkoinen miljöö vaihtelee lämpötilaltaan, kosteudeltaan ja ravintoaineitten saatavuudelta.
Hyvin tärkeä näkökohta homeostaasin ylläpidossa on kyky tunnistaa ”ystävä vihollisesta”, ” to recognize friend from foe”. Sen takia kaikki kehon solut kantavat identiteettitunnuksia, merkitsijöitä, osoittaakseen, että ne kuuluvat a.o. kehoon ja sellaiset solut, jotka eivät pysty identifioimaan "itseä", tuhotaan heti immuunijärjestelmän patrulloivilla ja vartioivilla soluilla.
Yksi uhka homeostaasille on kehoon invasoituvat patogeenit kuten virukset ja bakteerit. Toinen uhka on sellaiset solut, jotka alkavat kasvaa ilman rajoituksia ja aiheuttavat pahanlaatuisia kasvaimia. Mikä tahansa on uhkan luonne, immuunijärjestelmälla on vaste kuitenkin samantyylistä. Immuunijärjestelmän strategiana on pikaisesti tuhota hyökkääjä ja jos tämä epäonnistuu, ainakin rajoittaa infektion määrää. Esim. ihmisellä voi olla piilevä tuberkuloosi-infektio, joka pysyy paikallisen kontrollin alaisena, ellei henkilö tule heikompaan kuntoon tai iän mukana. Samoin premaligneja soluja voi keho tuhota tai pakottaa niitä ”vanhentumaan” ja häviämään. Jotta immuunijärjestelmä saisi maksimaalisen tehonsa, sitä koordinoi AIVO, keskushermosto (CNS), joka pystyy kehkeyttämään sairauden tuntemukset ja oireet, kuten KUUMEEN, ANOREXIAN ja VÄSYMYKSEN. AIVO, CNS taltioi myös muistiaineksena niitä kehon ottamia aineksia, jotka ovat aiemmin tehneet kehon sairaaksi ja saattaa henkilön välttämään sellaisen aistimukseen uudestaan joutumista ehdollisella makuaistimuksen aversiolla ( conditional taste aversion). (Huom, kommenttini: Tämä AIVO vaikuttaa ihmisindividin intelligenssiin ja tietoiseen minään ja persoonaan tunteita luomalla, joitten kautta CNS , varsinainen biologinen AIVO, säätelee elämälle edullisia päätöksiä kehossa asustavalta individiltä, joka harvoin valitsee elintapoja, jotka ovat AIVOLLE edullisimmat, joten varsinainen biologinen AIVO on jatkuvassa katastrofivalmiudessa inidividuellin impulsiivisen kekseliäisyyden takia).
Aktiivit immuunisolut vapauttavat SYTOKIINEJÄ, jotka antavat keskushermostolle signalointia. SYTOKIINIT vaikuttavat vapaisiin hermopäätteisiin, veri-aivoesteeseen (BBB, blood- brain- barrier) tai sellaisiin keskushermoston osiin, joista puuttuu BBB (veriaivoeste) ja ne saavat aikaan kuumetta, anoreksiaa ja väsymystä. Nämä oireet toimivat tehokkaina puolustuskeinoina lyhyen ajan sairauksissa auttamalla voittamaan patogeeniä, siten että kehon energiareservit saattavat keskittyä immuunisysteemin palveluksiin ja luoda sisäistä miljöötä, mikä on invasoituvalle patogeenille vähemmän soveltuva ja AIVO saa mahdollisuuksia toipua.
Mutta jos tähän patogeenin voittamiseen menee paljon aikaa, aineenvaihdunnan kohoaminen ja alentunut ravinnon otto voi koitua haitaksi yksilölle ja johtaakin anoreksia-kakeksia oireyhtymään, mitä havaitaan vakavissa kroonisssa taudeissa, kuten silloin, kun jokin elin menee vajaatoimintaan, kun infektio pitkittyy tai syöpä on alkanut. Niissä tiloissakin AIVO on hyvin pitkälle suojattu energeettisesti.
KAKEKSIA ( Cachexia)- tilassa on aineenvaihdunta kiihtynyt, painoa menetetään, anemiaa ja väsymystä esiintyy ja elinikä vaikuttuu, muita tauteja alkaa ilmetä samanaikaisesti ja henkilön vaivat enenevät.
Toinen näkökohta HOMEOSTAASISTA
Homeostaasi pitää yllä kudosten integriteettiä. Milloin tahansa ihon muodostama este ( skin barrier) särkyy tai jokin kehon kudos vaurioituu, vapaita hermopäätteitä (nociceptors), kivun aistijoita, alkaa olla signaloimassa aivoille, että aivojen on sopeutettava käyttäytyminen sellaiseksi, mikä vähentää vauriota. AIVOJEN funktio ilmenee käyttäytymisenä (behavior). Kivun aistiminen (nociception) on vahva motivoidun käyttäytymisen modulaattori ja nosiseptiset stimulukset aktivoivat autonomisen hermoston (ANS, Autonomic Nervous System) sympaattiset ( symphatethetic) säikeet. Sekä immuunijärjestelmä että nosiseptiivinen järjestelmä toimivat konjunktiossa keskenään. ESIM: tulehtuneessa kudoksessa nosiseptorit herkistyvät tulehduksen välittäjille ja tämä ilmiö tunnetaan hyperalgesiana, kohoneena kipuherkkyytenä.
Bakteerin lipopolysakkaridi LPS
LPS sitoutuu kehossa plasmassa kulkevaan proteiiniin LBP ( Lipopolysaccharide binding protein) ja se voi aktivoida immuunisoluja ja muodostaa kompleksin CD14 ( joka on plasmamebraaniin sitoutunut reseptoriproteiini). CD14 ei kuitenkaan omaa solun sisällä mitään jatketta, mikä informoisi solua tilanteesta, mutta signaalin edistyminen sai selityksensä aikanaan, kun keksittiin, että LPS voi tehdä interaktion immuunijärjestelmän kanssa. Löydettiin nimittäin Tollin reseptorit, joista lipoproteiineille ( lp) reagoi TLR2. sRNA ja dsRNA viruksia tunnistaa TLR3 ja TLR7. TLR4 tunnistaa LPS, bakteereitten lipopolysakkarideja. TLR5 tunnistaa flagelliinia. TLR9 tunnistaa CpG DNA.ta. Nykyinen ymmärtämys LPS- substanssin stimuloimasta immuunivasteesta on seuraava: Ensin akuutin faasin proteiini LBP tunnistaa sitä ja esittää immuunisolujen CD14-molekyylille. CD14 lataa sitä gpMD-2 glykoproteiiniin, joka voi muodostaa sitten TLR4 Tollin resptorin kanssa kompleksin. Tästä aktivoituu lopulta solun sisäinen tie adaptoriproteiinilla MyD88, joka siten rekrytoi seriinitreoniinikinaaseta, joita sanotaan IRAKs ja aktivoi TRAF6. Tästä taas aktivoituu transkriptiofaktoria NF-kB tai fos/jun mistä seuraa geenitranskriptiot seuraaville sytokiineille ja välittäjäaineille: TNFalfa, IL-18, COX-2, IL-1, IL-6 ja IL-8. ESIM. KUVA TLR4 kautta tapahtuvasta LPS aktivoimasta tiestä.
KUVA 6. -7.
http://www.iab.keio.ac.jp/ja/tlr-chart.jpg
{kind=link}
http://www.autoimmunityresearch.org/lyme-disease/image23.gif
{kind=link}
Tumassa sitten fos/jun, NF-kB transkriptiotekijöitten aktivaatio johtaa monien pro-inflammamtoristen sytokiinien geenien koodaamien ohjelmien esiinkirjoittamiseen ( transkriptio mRNA muotoon) ja mRNA taas voidaan translatoida, kirjoittaa peptidiksi tai proteiiniksi solun endoplasmisessa retikulumissa(ER) ja saadaan yllämainittuja proinflammatorisisa tuotteita.
Muistiin 2007-04-09
Päivitystä 10.2. 2011
Aivoille funktionaalista ruokaa on tavallinen ruoka. Aivojen toiminnalle on suotuista noudattaa raqvintosuosituksia energiasta ja ravintoaineista ja nämä suositukset ovat tehty AIVON taholta, eli yhteiskunnan korkeimman tieteellisen tiedon mukaan.
Kts ravinto ja energia-aineet.
http://pikenruoka.blogspot.com/
Inga kommentarer:
Skicka en kommentar